




Este artigo tenta explicar de maneira simples o que é a Biofísica, suas fundamentações e teorias, a respeito de uma ferramenta que amplia a visão da ciência como um leque de explicações sobre o funcionamento de organismos vivos, fazendo um elo com a biologia e a física que por hora, pode parecer distante uma da outra e que vão se entrelaçando quando lemos esse artigo e botamos em funcionamento a máquina que é o corpo humano.
A biofísica tem como característica o estudo da Biologia usando métodos e princípios científicos da Física. Podemos considerar que a ciência não orbita suas disciplinas marcantes como a química, biologia, e física, em órbitas separadas, e sim como um conjunto interdisciplinar.Estudar a Física aplicado nos seres vivos, pode ser muito útil em áreas como a Medicina, Biomedicina, Educação Física, Fisioterapia por exemplo.
Sua área de atuação é vasta como vimos acima, sem falar que o estudante pode compreender vários fenômenos que fazem parte do seu corpo que diariamente se transforma e esta sempre repetindo seus ciclos vitais para vida. Aspectos como eletricidade, eletromagnetismo, mecânica, e física nuclear fazem parte da composição de vários fenômenos biológicos. Em óptica estudamos a óptica da visão que estuda o funcionamento do olho que focaliza objetos e produz uma imagem do mesmo nítida quando não possui algum tipo de anomalia.
Nesta unção da Biologia com a Física o compartilhamento de teorias possibilita criar novos horizontes científicos, tais como a bioacústica que estuda a propagação sonora em meios elásticos e sua recepção pelos animais, incluindo os humanos, os avanços dessa área em especifico possibilitou o desenvolvimento de aparelhos auditivos que trouxe um maior conforto para quem sofre de alguma deficiência no aparelho auditivo.
Uma outra frente de pesquisa que deriva da Biofísica é a biomecânica que estuda ou aplica a mecânica na Biologia, sendo muito utilizada em áreas como a Educação Física, Fisioterapia, enquanto ciência, ela visa tratar aspectos cinemáticos do movimento humano ou seja o movimento mecânico dos sistemas vivos. A bioeletricidade esta relacionada com a voltagem estática das células bem como a corrente elétrica que flui através de nervos e músculos proveniente de um potencial de ação.
Fontes:
HALLIDAY, David, RESNIK Robert, KRANE, Denneth S. Física 4, volume 2, 5 Ed. Rio de Janeiro: LTC, 2004. 392 p.
HALLIDAY, David, RESNIK Robert, KRANE, Denneth S. Física 4, volume 2, 5 Ed. Rio de Janeiro: LTC, 2004. 392 p.
R. Glaser, Biophysics, Springer Verlag, Heidelberg, 1996.
E. Okuno, L, Caldas, C. Chow, Física para Ciências Biológicas e Biomédicas, Harper & Row do Brasil, São Paulo, 1982.
http://pt.wikipedia.org/wiki/Bioeletricidade
No corpo humano - a fonte de energia é o alimento; ele é processado no sistema digestivo e é então combinado com O2 nas células do corpo para liberar energia. Seus subprodutos são expelidos de 4 formas:
as componentes não digestivas são eliminadas como fezes (liberando somente uma pequena quantidade de gases nocivos),
água e outros subprodutos são expelidos pela urina,
é expelido via pulmões a cada dia, e
o calor é dissipado pela superfície do corpo.
água e outros subprodutos são expelidos pela urina,
é expelido via pulmões a cada dia, e
o calor é dissipado pela superfície do corpo.
A “máquina” humana realmente consiste de bilhões de “máquinas” muito pequenas – as células vivas do corpo. Cada uma destas “máquinas – miniaturas” deve ser provida com combustível, O2, e um método de livrar-se dos subprodutos. O sangue e seus vasos (o sistema cardiovascular) serve como sistema de transporte para estas máquinas. Os pulmões (o sistema pulmonar) serve como fornecedor de O2 e expelidor do principal subproduto – CO2. O sangue leva o O2 aos tecidos e remove o CO2 dos tecidos; ele deve entrar em estreito contato com o ar nos pulmões a fim de trocar sua carga de CO2 por uma carga fresca de O2 (Fig.1). Os detalhes deste processo são discutidos na seção.2.
Por causa da estreita cooperação e interação entre os sistemas pulmonar e cardiovascular, as ações de um sistema freqüentemente afetam o outro. Por exemplo, durante a respiração a pressão nas veias principais do peito afetam o retorno do sangue ao coração. Freqüentemente uma doença dos pulmões produzirá sintomas no coração e vice-versa
Por causa da estreita cooperação e interação entre os sistemas pulmonar e cardiovascular, as ações de um sistema freqüentemente afetam o outro. Por exemplo, durante a respiração a pressão nas veias principais do peito afetam o retorno do sangue ao coração. Freqüentemente uma doença dos pulmões produzirá sintomas no coração e vice-versa
Figura 7.1 Os canais principais para o ar e sangue no sistema respiratório. O diagrama ilustra esquematicamente as trocas de O2 e CO2 entre o ar nos alvéolos e o sangue. (De Guyton, A.C., Function of the Human Body, 3rd ed. , Ó W.B. Saunders Company, Philadelphia, 1969, p. 222, Reprinted by permission.)
Os pulmões realizam outras funções fisiológicas além das trocas de O2 e CO2. Uma função principal é manter o pH (acidez) do sangue constante. Os pulmões desempenham um papel secundário nas trocas de calor (ver Capítulo 5) e o balanço fluido do corpo por aquecimento e mistura do ar que respiramos (inspiramos). Nosso mecanismo de respiração produz um fluxo controlado de ar para falarmos, tossirmos, espirrarmos, suspirarmos, soluçarmos, sorrirmos, fungarmos e bocejarmos. Em adição, bloqueando a passagem de ar gera aumentos de pressão para defecar e vomitar.
Uma importante função do aparelho respiratório é a produção da voz. As formas de respiração são marcadamente diferentes durante a conversação. Desde que a voz é produzida por uma saída controlada de ar dos pulmões, uma pessoa inala rapidamente e muito profundamente antes de falar afim de ter mais tempo para produzir sons vocais. O tempo de inalação é tipicamente menos de 20% do ciclo da respiração, e a quantidade inalada é geralmente mais que duas vezes o volume usual. Uma cantora (especialmente uma cantora de ópera) inala mesmo mais ar num curto período de tempo para posteriormente minimizar as partes inalatórias do ciclo. A resistência aérea produzida pelas cordas vocais causam um aumento na pressão relativamente grande na traquéia. Assim, o trabalho envolvido no falar e no cantar é consideravelmente maior que o trabalho da respiração normal. Entretanto, relativamente pouco do aumento de trabalho participa da energia sonora.
A voz tipicamente tem uma potência menor que 1 mW. A produção da voz será discutida posteriormente.
Nós respiramos cerca de 6 litros de ar por minuto. (Isto é também cerca do volume de sangue que o coração bombeia cada minuto). A respiração do homem em repouso é cerca de 12 vezes por minuto, a mulher respira cerca de 20 vezes por minuto, e as crianças respiram cerca de 60 vezes por minuto. Discutimos na Seção 7.6 os fatores físicos que afetam a razão de respiração.
O ar que nós inspiramos é cerca de 80% de N2 e 20% de O2. O ar expirado é cerca de 80% de N2, 16% de O2 e 4% de CO2. Nós respiramos cerca de 10 kg (22 lb) de ar a cada dia. Disto o pulmão absorve cerca de 400 litros (~ 0,5 kg) de O2 e libera uma quantidade ligeiramente menor de CO2. Nós também saturamos o ar que nós respiramos com água. Quando respiramos ar seco, nosso ar expirado leva para fora cerca de 0,5 kg de água a cada dia. (Esta mistura pode ser usada para limpar os óculos). No frio alguma desta mistura se condensa e vemos a nossa respiração.
O ar que nós respiramos contém poeira, fumaça, carga de bactérias, gases nocivos e assim por diante, que entram em contato íntimo com o sangue. A grande superfície torcida dos pulmões com uma área superficial de cerca de 80 m2tem uma exposição maior ao ambiente que qualquer outra parte do corpo incluindo a pele. É talvez surpreendente nós não termos mais doenças dos pulmões. A importância de ar limpo é óbvia.
Cada vez que nós respiramos, cerca de 1022 moléculas de ar entram em nossos pulmões. Lembrem-se que 22,4 litros de ar contém cerca de 6 x 1023 moléculas – número de Avogadro. O número total de moléculas na atmosfera terrestre é cerca de 1044. Assim, tomamos 1/1022 de todo o ar terrestre a cada vez que respiramos; em outras palavras, para cada molécula que respiramos existem 1022 outras mais na atmosfera. A atmosfera terrestre está em constante movimento e sobre um período de séculos tem existido através de mistura de gases. Como um resultado, cada 0,5 litros de ar (1022 moléculas ) contém em média uma molécula que estava presente em qualquer 0,5 litro de ar séculos atrás. Um modo interessante de pensar sobre isto é que na média cada uma de nossa respiração contém uma molécula de ar que foi respirada por Cristo em qualquer uma de suas respirações. É claro, isto é verdadeiro para qualquer pessoa, famosa, não famosa, ou desconhecida que viveu muitos anos atrás.
Problema 1 - Calcule o número de moléculas de O2 absorvidas pelo corpo numa respiração típica de 500 cm3.
AS VIAS AÉREAS
Diagrama esquemático mostrando as principais passagens de ar para os pulmões
As principais passagens aéreas para os pulmões estão mostradas na Fig 7.2. O ar entra normalmente no corpo através do nariz onde ele é aquecido (se necessário), filtrado e misturado. As superfícies úmidas e os pêlos no nariz aprisionam particles de poeira, micróbios, e assim por diante. Durante exercícios pesados, tais como jogando, o ar é respirado através da boca e escapam deste sistema de filtro. O ar então passa através da traquéia. A traquéia se divide em duas (bifurca) para fornecer o ar a cada pulmão através dos brônquios. Cada brônquio se divide e subdivide cerca de 15 vezes mais; os bronquíolos terminais resultantes alimentam de ar os milhões de pequenos sacos chamados alvéolos. Os alvéolos, que são como pequenos balões interconectados (Fig. 7.3), têm mais ou menos 0,2 mm de diâmetro (uma folha de papel é aproximadamente 0,1 mm de espessura) e tem paredes de somente 0,4 m m de espessura. Eles expandem-se e contraem-se durante a respiração; eles estão “onde a ação está” na troca de O2 e CO2. Cada alvéolo é envolvido por sangue de modo que O2 possa difundir dos alvéolos para os glóbulos vermelhos e CO2 possa difundir do sangue para o ar no alvéolo. No nascimento os pulmões tem cerca de 30 milhões de alvéolos; aos 8 anos de idade o número de alvéolos cresce para cerca de 300 milhões. Além desta idade o número permanece relativamente constante, mas o alvéolo aumenta de diâmetro. Os alvéolos desempenham um papel tão importante na respiração que discutiremos a física dos alvéolos com mais detalhes na seção 7.5.
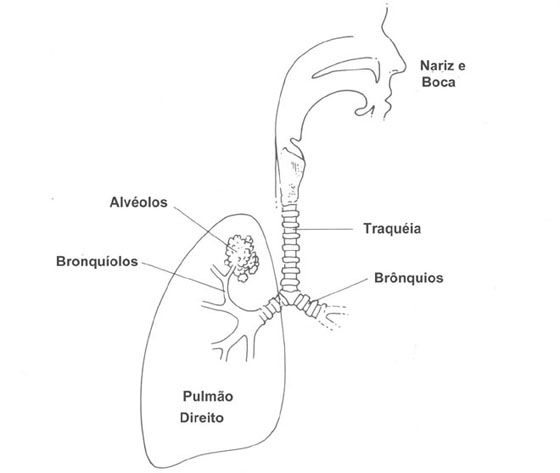
Figura 7.2 - Diagrama esquemático mostrando as passagens principais de ar para os pulmões.
Afim de servir como sistema de transporte para o ar, as vias respiratórias removem as partículas “poeira” que atingem a úmidas vias das várias passagens de ar. O corpo tem dois mecanismos para limpeza das vias respiratórias de partículas estranhas. Os mais corpulentos são removidos pelo tosse. Partículas pequenas são levadas de cima para baixo para a boca por milhões de pequenos pêlos, ou álios. Os álios, que são somente cerca de 0,1 mm de comprimento, tem um movimento ondulatório que move mucosa levando poeira e outras pequenas partículas até os vias respiratórias maiores. Cada um dos álios vibram cerca de 1 000 vezes por minuto. A mucosa move cerca de 1 a 2 cm/min. Você pode pensar nos álios como um sistema de escada rolante para a traquéia. Leva cerca de 30 min para uma partícula de poeira ser retirada dos brônquios e da traquéia para a garganta onde ela é expelida ou engolida.
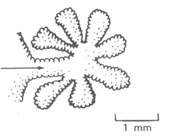
Figura 7.3 – A estrutura do alvéolo
7.2 – COMO O SANGUE E OS PULMÕES INTERAGEM
O principal propósito da respiração é levar um suprimento de O2 fresco para o sangue nos pulmões e desfazer-se do CO2. Nesta seção tentaremos ajudá-lo a entender a física envolvida na troca de gás entre os pulmões e o sangue.
O sangue é bombeado do coração para os pulmões sob pressão relativamente baixa. A pressão sangüínea média de pico na principal artéria pulmonar que leva o sangue aos pulmões é somente cerca de 20 mmHg ou cerca de 15% da pressão na circulação principal do corpo. Os pulmões oferecem pouca resistência ao fluxo de sangue. Na média, cerca de um quinto (~ 1 litro) do sangue que alimenta o corpo está nos pulmões, mas somente cerca de 70 ml daquele sangue está nos capilares dos pulmões obtendo O2 em qualquer instante. Desde que o sangue fica nos capilares pulmonares por menos de 1 segundo, os pulmões devem ser bem projetados para trocas gasosas; os alvéolos dos pulmões tem paredes extremamente finas e são envolvidos pelo sangue no sistema capilar pulmonar. A área da superfície entre o ar e sangue nos pulmões é cerca de 80 m2 (cerca da metade da área de uma quadra de tênis). Se os 70 ml de sangue nos capilares pulmonares forem espalhada sobre uma área superficial de 80 m2 a camada resultante de sangue será somente cerca de 1 mm de espessura, menos do que a espessura de uma única célula de glóbulo vermelho.
Dois processos gerais estão envolvidos nas trocas gasosas nos pulmões:
Se um ou outro processo falha o sangue não será oxigenado apropriadamente.
Existem três tipos de áreas aspersão-ventilação nos pulmões:
Num pulmão normal o primeiro tipo ocorre com mais de 90% do volume total. Se o sangue que flui para uma parte de um pulmão é bloqueado por um coágulo (uma embolia pulmonar) aquele volume terá aspersão pobre. Se as passagens aéreas nos pulmões estão obstruídas como na pneumonia, a área envolvida terá ventilação pobre. Muitas doenças pulmonares causam reduções na aspersão ou ventilação.
A transferência de O2 e CO2 para dentro e para fora do sangue é controlada pela lei física da difusão. Todas as moléculas estão continuamente em movimento. Nos gases e nos líquidos, e numa certa extensão mesmo nos sólidos, as moléculas não permanecem numa determinada localização. Por exemplo, se você pode identificar um grupo de moléculas numa sala (p. ex., de uma gota de perfume) em poucos minutos você encontrará que estas moléculas moveram-se (difundiram-se) pela sala. Moléculas de um tipo particular difundem-se de uma região de alta concentração para uma região de concentração mais baixa até que a concentração seja uniforme. Nos pulmões estamos interessados na difusão de gases e líquidos. As moléculas num gás na temperatura da sala movem-se aproximadamente com a velocidade do som. Cada molécula colide cerca de 1010 vezes em cada segundo com as moléculas vizinhas, peregrinando no processo de uma maneira aleatória. A distância D mais provável que uma molécula viajará de sua origem após N colisões é D = l EXEMPLO 7.1
Qual é o valor típico de D no ar e no tecido para uma molécula de O2 após 1 segundo se N = 1010 no ar e N = 1012 no tecido?
No ar D ~ 10-7 (1010)1/2 = 10-2 m
No tecido D ~ 10-11 (1012)1/2 = 10-5 m.
A difusão depende da velocidade das moléculas; ela é mais rápida se as moléculas são leves e ela cresce com a temperatura. Desde que N é proporcional ao tempo de difusão Dt (i.é, N a Dt), podemos escrever que D a 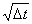 ouDt a D2. Se D = 10 mm após 1 segundo, a molécula levará 100 segundos, em média, para difundir 100 mm. Nos pulmões a distância a ser viajada no ar é usualmente uma pequena fração de um milímetro, e a difusão toma lugar numa fração de um segundo. A difusão de O2 e CO2 no tecido é cerca de 10 000 vezes mais lenta que no ar, mas a espessura do0tecido pelo qual a molécula deve difundir-se nos pulmões é muito pequena (~0,4 mm ) e a difusëo através di parede alveolar leva muito menos que 1 segundo. Discutimos mais sobre a difusão nos tecidos em outro Apêndice.
ouDt a D2. Se D = 10 mm após 1 segundo, a molécula levará 100 segundos, em média, para difundir 100 mm. Nos pulmões a distância a ser viajada no ar é usualmente uma pequena fração de um milímetro, e a difusão toma lugar numa fração de um segundo. A difusão de O2 e CO2 no tecido é cerca de 10 000 vezes mais lenta que no ar, mas a espessura do0tecido pelo qual a molécula deve difundir-se nos pulmões é muito pequena (~0,4 mm ) e a difusëo através di parede alveolar leva muito menos que 1 segundo. Discutimos mais sobre a difusão nos tecidos em outro Apêndice.
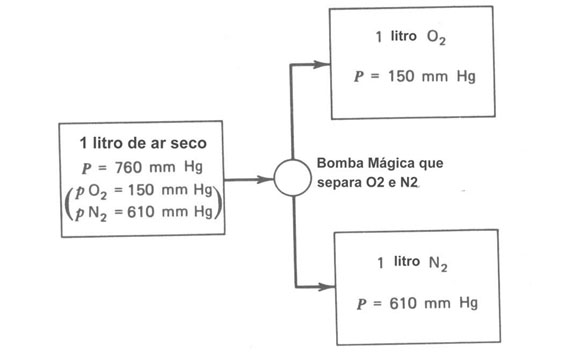
Para entender o comportamento dos gases nos pulmões é necessário revisar a lei de Dalton das pressões parciais. Esta lei diz que se você tem uma mistura de vários gases, cada gás faz a sua própria contribuição à pressão total imaginada como se ele fosse o único. Consideremos um recipiente fechado de ar seco à pressão atmosférica (760 mmHg). Se você removeu do recipiente todas as moléculas exceto O2, a pressão cairá para cerca de 150 mmHg (i.é, 20% de 760 mmHg). Esta é a pressão parcial do oxigênio pO2. Se somente as moléculas de N2 forem deixadas, a pressão será cerca de 80% de 760 mmHg ou cerca de 610 mmHg. A Figura 7.4 mostra esquematicamente este experimento imaginário. A pressão parcial do vapor d’água no ar depende da umidade. No ar de uma sala típica, a pressão parcial do vapor d’água é de 15 a 20 mmHg; nos pulmões a 37ºC e 100% de umidade relativa a pressão parcial do vapor d’água é 47 mmHw.
Consideremos o que acontece num recipiente fechado com sangue e O2. Algumas moléculas de O2 colidiróo com o sangue e serão dissolvidas. Após, um tempo o número de molésulas de O2 que estão(escapando do sangue a cada segundo é o mesmo número das que(estão entraodo nele/ O sangue tem então uma pO2 igual aquela do O2 em contato com ele. Se a pO2 na fase gasosa é eobrada, a quantidade de O2 dissolvida no sangue também será dobrada. Esta proporcionalidade é a lei de Henry da solubilidade dos!gases.
A quantidade de gás disóolvida no sangue varia castante de um gás para o outro. O oxigênio não é muito solúvel oo sangue ou água. À temperaôura(do corpo, um litro de plasma saogüíneo à pO3 te 100 mmHg manterá somente cerca de 2,5 cm3 de O2 a temperatura e pressão normal (C.N.T.P.). Em pO2 de 40 mmHg manterá cerca de 25 cm3 de CO2 na solução. Se o corpo teve de depender de O2 dissolvido no plasma para suprir O2 para as células, o coração terá que bombear 140 litros de sangue por minuto no repouso, em vez dos 6 litros/minuto que ele realmente bombeia. Como discutimos brevemente, existe um método mais eficiente de transporte de O2 e CO2 que envolve os glóbulos vermelhos.
As diferentes solubilidades de O2 e CO2 no tecido afetam o transporte destes gases através da parede alveolar. Uma molécula de O2 difunde-se mais rapidamente que uma molécula de CO2 devido a sua massa ser menor. Entretanto, por causa do maior número de moléculas de CO2 na solução, o transporte de CO2 é mais eficiente que o transporte de O2. Se uma doença torna a parede alveolar mais espessa, o transporte de O2 é mais prejudicado que o transporte de CO2.
A mistura de gases nos alvéolos não é a mesma que a mistura de gases no ar ordinário. Os pulmões não são esvaziados durante a expiração. Durante a respiração normal os pulmões retém cerca de 30% do seu volume no final de cada expiração. Isto é chamado “CAPACIDADE RESIDUAL FUNCIONAL” (F.R.C.). Em cada respiração cerca de 500 cm3 de ar fresco (pO2 de 150 mmHg) mistura-se com cerca de 2 000 cm3 de ar viciado nos pulmões para resultar no ar alveolar com uma pO2 de cerca de 100 mmHg. O pCO2 nos alvéolos é cerca de 40 mmHg. O ar expirado inclui cerca de 150 cm3 de ar relativamente fresco da traquéia que não estava em contato com as superfícies alveolares, assim o ar expirado tem uma pO2 ligeiramente mais alta e uma pCO2 mais baixa que o ar alveolar (Tabela 7.1). A razão da saída do CO2 pela entrada de O2 é chamada RAZÃO DE TROCA RESPIRATÓRIA ou QUOCIENTE RESPIRATÓRIO R (ver Capítulo 5). Geralmente R é ligeiramente menor que 1.
As porcentagens e Pressões Parciais de O2 e CO2 no Ar Inspirado, Alveolar e Expirado
O nitrogênio do ar não desempenha qualquer papel conhecido nas funções do corpo. Ele é dissolvido no sangue na sua pressão parcial. Um mergulhador no mar respira ar numa pressão muito mais alta submerso do que quando ele está no nível do mar; a pressão parcial aumentada de N2 faz com que mais N2 seja dissolvido no seu sangue e tecidos. Se o mergulhador vem à superfície muito rapidamente alguns dos N2 formam bolhas nas suas juntas causando os sérios problemas de cãimbras (ver Capítulo 6).
Durante a respiração normal o suprimento de ar fresco não entra nos alvéolos que estão ainda cheios de ar viciado da respiração anterior. Por causa da sua maior concentração, o O2 fresco rapidamente difunde pelo ar viciado até atingir a superfície dos alvéolos. O O2 é dissolvido na mistura parede alveolar e difunde por ela até o capilar sangüíneo até que a pO2 no sangue seja igual aquela dos alvéolos. Este processo leva menos do que 0,5 segundos (Fig. 7.5). Entretanto, o CO2 no sangue difunde-se mesmo mais rapidamente no gás dos alvéolos até que a pCO2 no sangue seja a mesma que no gás alveolar.
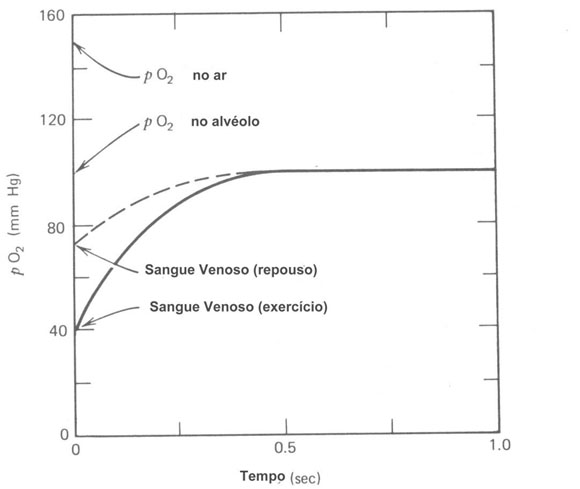
Como mencionado anteriormente, o sangue pode carregar muito pouco O2 na solução. A maioria do O2 para as células é carregado na combinação química com a hemoglobina (Hb) nos glóbulos vermelhos. Um litro de sangue pode carregar cerca de 200 cm3 de O2 nas C.N.T.P., por este meio enquanto ele pode carregar somente 2,5 cm3 de O2 em solução. Desde que a maioria do O2 não está em solução, a lei de difusão á alterada. O O2 combinará com ou separará da Hb de uma maneira que depende da curva de dissociação (Fig. 7.6). A Hb que deixa os pulmões está cerca de 97% saturada com O2 numa pO2 de cerca de 100 mmHg. O pO2 tem que cair para cerca de 50% antes da carga de O2 do sangue é notavelmente reduzida.
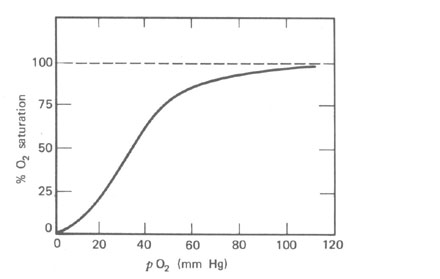
Quando o sangue atinge as células e seu baixo pO2 ambiente, o O2 é dissociado do Hb e difunde na célula. Nem todo o O2 deixa a Hb; a quantidade deixada depende do pO2 dos tecidos. Sob condições de repouso o sangue venoso retorna ao coração com cerca de 75% da sua carga de O2. O O2 é retido no sangue porque não é necessário nos tecidos. Durante trabalhos físicos pesados ou exercícios a situação nos músculos ativos variam drasticamente. O pO2nos músculos em funcionamento cai rapidamente fazendo mais O2 ser dissociados da Hb e difundir para o interior dos músculos. Em adição o corpo pode aumentar o fluxo sangüíneo para o trabalho muscular por um fator de 3. Os músculos trabalhando podem obter 10 vezes mais O2 que eles consomem no repouso. Para pessoas normais o fator limitante no exercício não é a quantidade de sangue bombeado pelo coração por minuto (saída cardíaca) ou a quantidade de O2 fornecida ao sangue pelos pulmões, mas a velocidade em que O2 é transferido aos músculos trabalhando.
A dissociação de O2 da Hb também depende do pO2, do pH (acidez) e da temperatura. Durante os exercícios o pCO2, a acidez e a temperatura nos músculos trabalhando aumentam todos; este aumento desloca todas as curvas da Fig. 7.6 para a direita e permite a Hb dar mais do seu O2. Todos estes fatores aumentam assim o O2 para os músculos trabalharem. Nos pulmões a diminuição de pCO2 devido a rápida respiração permite Hb agarrar mais O2.
Dióxido de carbono não é transportado dos tecidos por simples difusão. (Se você está interessado nos detalhes ver bibliografia no final deste capítulo). A maioria do CO2 permanece no sangue após ele ter deixado os pulmões (pCO2~ 40 mmHg). O nível de CO2 no sangue é mantido bem constante pela razão de respiração. Respiração excessivamente rápida (hiperventilação) pode abaixar o pCO2 do sangue (hipocapnia); isto provoca distúrbios mentais e desmaios.
No envenenamento por monóxido de carbono (CO) moléculas são grudadas muito seguramente à Hb nos lugares que normalmente seriam ocupados por O2. Elas grudam cerca de 250 vezes mais que o O2 e não se dissociam facilmente nos tecidos. Em adição usando os lugares normalmente usados para transportar O2, o CO impede a liberação de O2 da Hb, todavia uma pequena quantidade de CO possa seriamente reduzir o O2 nos tecidos. Fumantes de cigarros respiram em cerca de 250 cm3 de CO em cada pacote, e também é geralmente inalado por pessoas viajando em tráfego pesado. Monóxido de carbono pode causar a morte por desnutrição dos tecidos de O2. Normalmente o O2 dissolvido no sangue é de não significado, mas se uma vítima de CO é colocada numa câmara hiperbárica de O2 com uma pressão absoluta de 3 atm de O2 puro, o pO2 aumenta por um fator de 15. O O2 dissolvido no sangue pode então suprir as necessidades mínimas do corpo (ver Capítulo 6). Esta terapia não pode ser mantida por muito tempo porque o envenenamento por O2 pode resultar. Uso continuado de 1 atm de O2 puro pode causar edema dos tecidos dos pulmões, que reduz o O2 no sangue e ironicamente resulta na morte de uma falta de O2 (anóxia = deficiência de oxigênio nos tecidos). Níveis de proteção de pO2 no “ar” são aqueles abaixo de 0,5 atm (~ 380 mmHg pO2) (ver Fig. 6.8).
7.3 MEDIDAS DOS VOLUMES PULMONARES
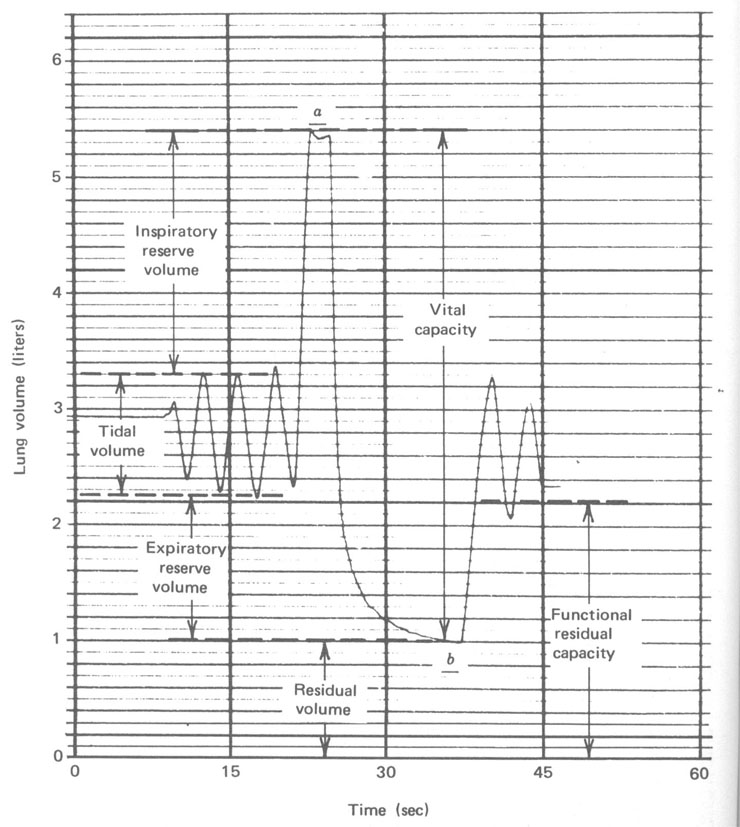
Um instrumento relativamente simples, o espirômetro (Fig. 7.7), é usado para medir o fluxo de ar para dentro e para fora dos pulmões e registrar num gráfico o volume versus o tempo. A Figura 7.8 mostra um registro típico para um adulto sob várias condições de respiração. Durante a respiração normal no repouso nós inalamos cerca de 500 cm3 de ar em cada respiração. Isto é referido como volume tidal (maré) no repouso. No começo e no final de uma respiração normal existe reserva considerável. No final de uma inspiração normal é possível com algum esforço promover o enchimento dos seus pulmões com ar. O ar adicional tomado é chamado volume de reserva inspiratório. Similarmente, no final de uma expiração normal você pode forçar mais ar para fora dos seus pulmões . Este ar adicional expirado é chamado volume de reserva expiratório. O ar restante nos pulmões após uma expiração normal é chamado capacidade residual funcional (FRC). É este ar velho que mistura com o ar fresco na nova respiração. Durante exercícios pesados, o volume tidal é consideravelmente maior. Você tem uma clara idéia da sua capacidade pulmonar se você tem mesmo soprar um saco de papel ou um colchão de ar. Se uma pessoa respira tão profundamente quanto pode (a em Fig. 7.8) e então exala tanto quanto pode (b em Fig. 7.8), o volume de ar exalado é a suacapacidade vital. Entretanto, seus pulmões ainda conterá algum ar – o volume residual, que é cerca de 1 litro para um adulto. O volume residual pode ser determinado tendo submetido a respiração num volume conhecido de um gás inerte tal como o hélio e então medindo a fração do hélio no gás expirado. Desde que o hélio e o ar misturar-se-ão durante uma única respiração, esta técnica de diluição é muito acurada.
Uma grande quantidade de testes clínicos podem ser feitos com o espirômetro. A quantidade de ar respirado em 1 minuto é chamado volume respiratório por minuto. O máximo volume de ar que pode ser respirado em 15 segundos é chamado ventilação voluntária máxima e é uma quantidade clínica útil . A máxima razão de expiração após uma inspiração máxima é um teste útil para enfisema e outras doenças das vias respiratórias obstruídas. Em alguns casos a razão de fluxo decresce com o esforço expiratório excessivo. Uma pessoa normal pode expirar cerca de 70% de sua capacidade vital em 0,5 segundos, 85% em 1,0 segundos, 94% em 2,0 segundos, e 97% em 3,0 segundos. Os picos normais da razão de fluxo são 350 a 500 litros/min. A velocidade do ar expirado pode ser impressiva; se uma pessoa engole ou espirra forte sem cobri sua boca, a velocidade do ar na sua traquéia pode atingir Mach 1 – a velocidade do som no ar! Esta velocidade alta pode causar colapso parcial das vias respiratórias por causa do efeito de Bernoulli. Engolindo para expulsar um objeto estranho este colapso parcial aumenta a velocidade do ar e aumenta a força no objeto estranho.
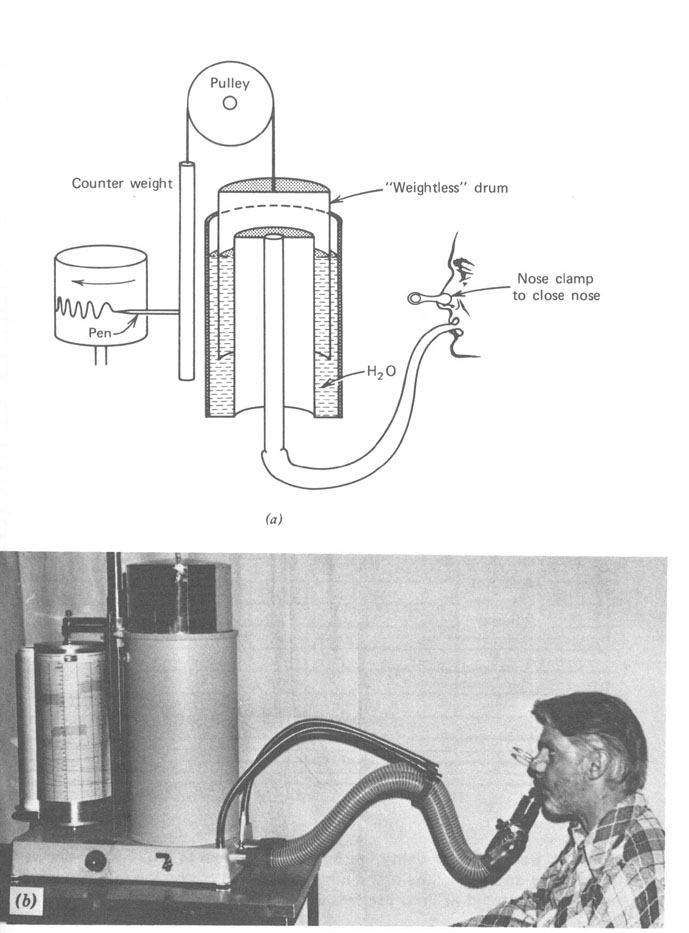
Figura 7.7 - O espirômetro é usado para medir várias quantidades de funçòes pulmonares. O fluxo de ar para dentro e para fora dos pulmões é registrado num cartão giratório. (a) Uma seção transversal do espirômetro mostrando como a água é usada como um selo de ar para manter o ar dentro do tambor contrabalançado. (b) Um dos autores (JRC) produzindo o gráfico mostrado na figura 7.8. O nariz tampado força todo o ar fluir através da boca
Nem todo o ar inspirado adiciona O2 ao sangue. O volume da traquéia e brônquios é chamado espaço anatômico morto já que o ar neste espaço não é exposto ao sangue nos capilares pulmonares. O espaço anatômico morto é cerca de 150 cm3. Em adição , em algumas doenças alguns dos capilares alveolares não são aspergidos com sangue e o O2 não é absorvido neste alvéolo. Este volume não usado é chamado fisiológico ou espaço alveolar morto. O ar no espaço alveolar morto não fornece qualquer O2 ao corpo. O ar no espaço anatômico morto após uma expiração é levado de volta para os pulmões durante a próxima inspiração. Se você aumentar o seu espaço morto respirando através de um tubo , você reciclará mais de sua própria respiração. Se o tubo tem um volume igual à sua capacidade vital você obviamente não obterá ar fresco e sufocará.
4 RELAÇÕES DA PRESSÃO-FLUXO DE AR-VOLUME DOS PULMÕES.
As relações de pressão, fluxo de ar, e volume dos pulmões durante a respiração tidal para um sujeito normal e para um paciente com uma estreita via aérea estão mostradas na Fig.7.9. A diferença de pressão necessária para fazer o ar fluir para dentro e para fora dos pulmões de um indivíduo saudável é muito pequena. Note que a diferença de pressão (Fig. 7.9a) é somente uns poucos centímetros de água para um indivíduo normal. A Figura 7.9b mostra a razão do fluxo de ar para dentro e para fora dos pulmões em litros por minutos, e a Fig. 7.9c mostra o volume dos pulmões durante o ciclo da respiração.
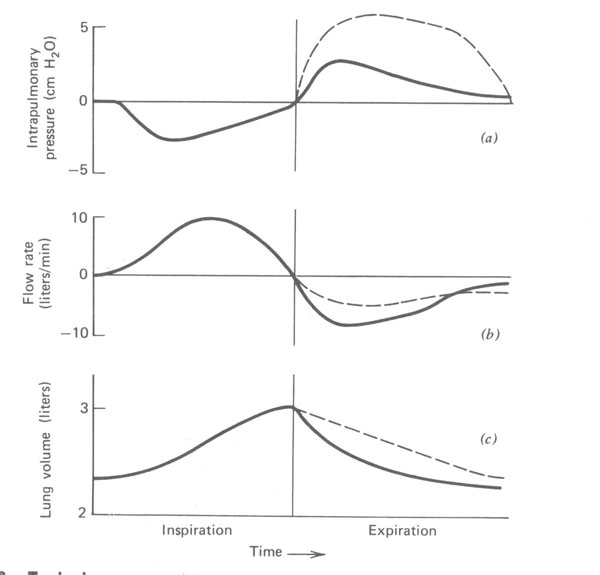
Figura 7.9 - Pressões típicas (a), razão de fluxo (b), e volumes pulmonares (c) durante uma respiração tranquila para um indivíduo normal (linha sólida) e um paciente com uma via respiratória estreita (linha tracejada). Note o acréscimo de pressão e o decréscimo da razão de fluxo durante a expiração no paciente com as vias respiratórias estreitadas
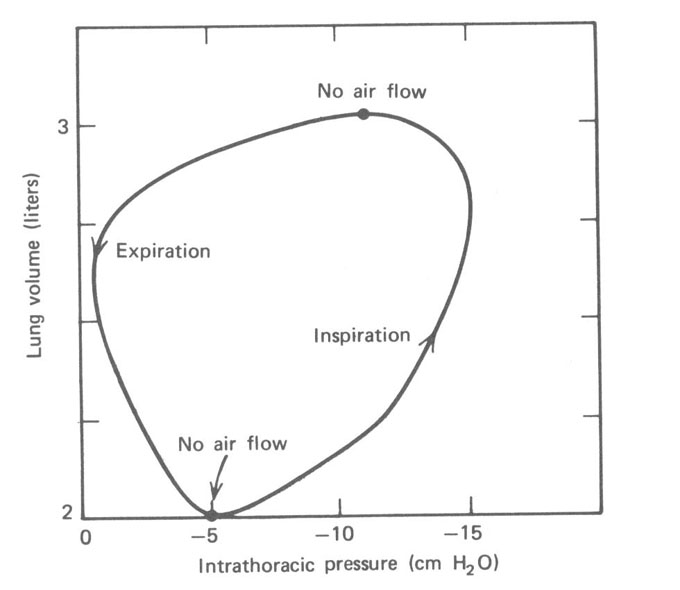
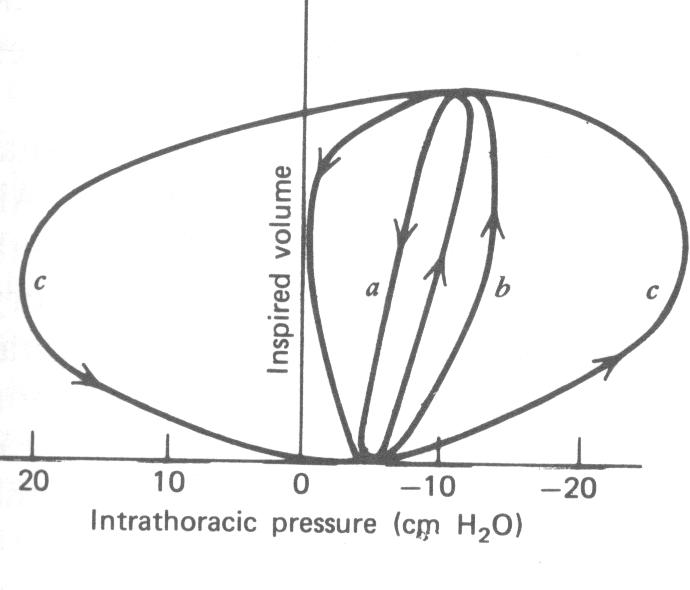
Como o esôfago passa através do peito, ele reflete a pressão entre os pulmões e a parede do peito (intrapleural ou espaço intratorácico). É possível medir a pressão no esôfago com uma pressão gauge. Esta pressão é normalmente negativa (~ -10 mmHg) devido à elasticidade dos pulmões (ver Seção 7.6). Na Fig. 7.10, a pressão intratorácica (medida no esôfago) é plotada versus o volume tidal dos pulmões durante a respiração. A Figura 7.11 mostra as curvas pressão-volume para três diferentes razão de respiração – lenta, moderada e rápida.
Os pulmões e o peito estão normalmente acoplados. O comportamento do sistema é o resultado da combinação das características físicas dos dois. A Figura 7.12 mostra as curvas de volume versus pressão para as paredes do peito e pulmões separadamente e para os dois juntos. O volume é dado como uma porcentagem da capacidade vital. Se a parede do peito estivesse livre de sua interação com os pulmões ela teria um volume de cerca de dois terços da capacidade vital total. Os pulmões por si só murchariam e não teriam volume de ar. Juntos os pulmões e a parede do peito vão a um volume de relaxação (FRC) de cerca de 30% de capacidade vital.
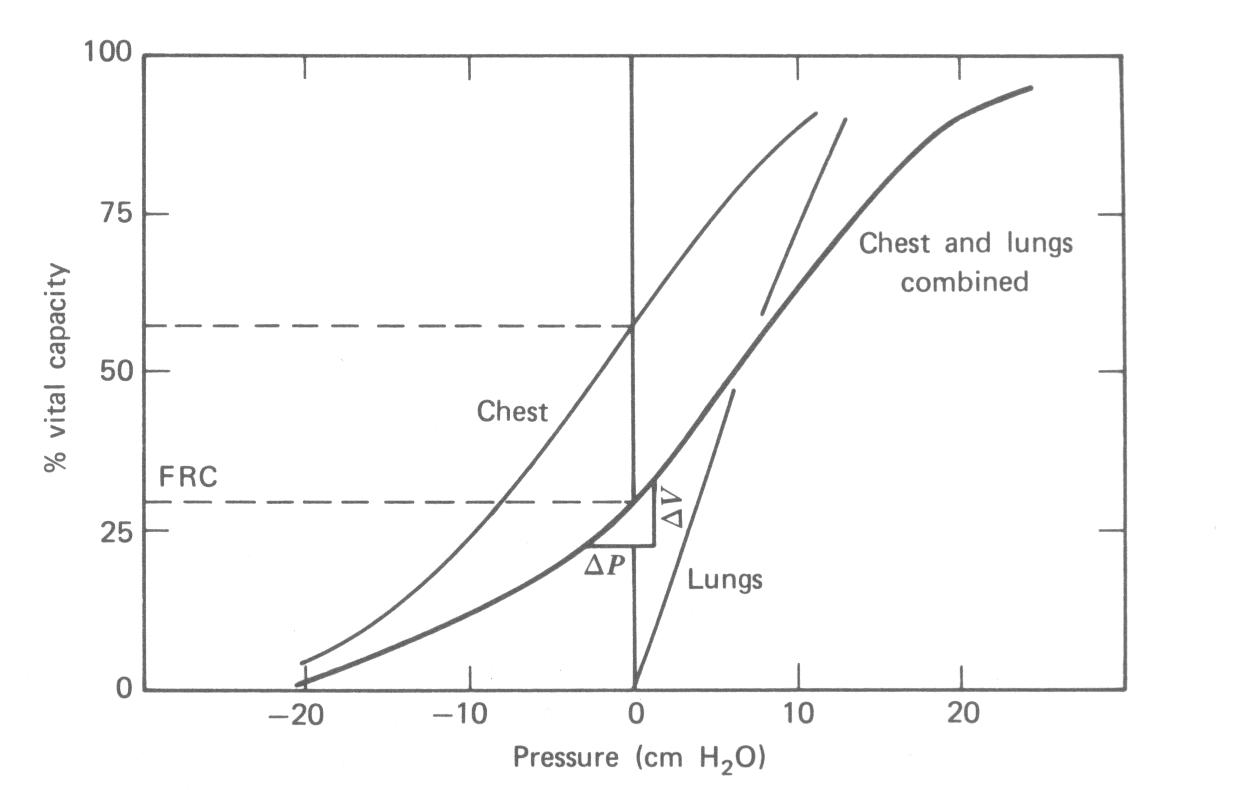
A curva combinada na Fig. 7.12 mostra a relação pressão-volume obtida enchendo os pulmões para conhecer as porcentagens da capacidade vital. A pressão é medida na boca (e pulmões) com o nariz e boca fechados e os músculos da respiração relaxados. Por exemplo, em cerca de 60% de capacidade vital, a pressão de relaxação é 10 cm H2O. Como a parede do peito está em equilíbrio neste volume, esta pressão é produzida pelas propriedades elásticas dos pulmões. Quando a mesma medida de relaxação são feitas após uma exalação forçada os valores de pressão negativa da Fig. 7.12 são obtidos.

0 comentários:
Postar um comentário